



For a hominoid fossil to be definitively classified as part of the human evolutionary line, certain evidence of bipedalism is required. Bipedalism is associated with anatomical changes literally from head to toe.
Bipedalism can even be preserved in the skull (Figure 7.1) because balancing the head in an upright posture requires a skull position relatively centered above the spinal column. The spinal cord leaves the skull at its base through an opening called the foramen magnum (Latin for “big opening”). In a knuckle-walker like a chimp, the foramen magnum is placed more toward the back of the skull while in a biped it is in a more forward position.
Extending down from the skull of a biped, the spinal column makes a series of convex and concave curves that together maintain the body in an upright posture by positioning the body’s center of gravity above the legs rather than forward. The curves correspond to the neck (cervical), chest (thoracic), lower back (lumbar), and pelvic (sacral) regions of the spine, respectively. In a chimp, the shape of the spine follows a single arching curve (Figure 7.2). Interestingly, at birth the spines of human babies have a single arching curve as seen in adult apes. As humans mature the curves characteristic of bipedalism appear, the cervical curve at about 3 months on average and the lumbar curve at around 12 months—a time when many babies begin to walk.

Fossilized footprints were preserved in volcanic ash at the 3.6-million-year-old Tanzanian site of Laetoli. As shown here, the foot of a living human fits right inside this ancient footprint, which shows the characteristic pattern of bipedal walking. The actual trail of footprints is 24 meters (80 feet) long.
Homo sapiens Australopithecus Ape
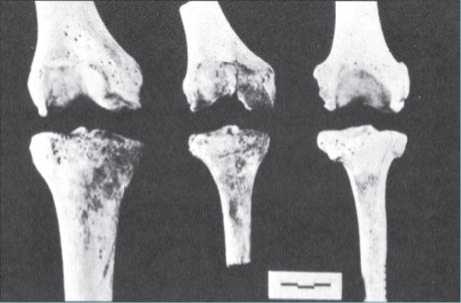
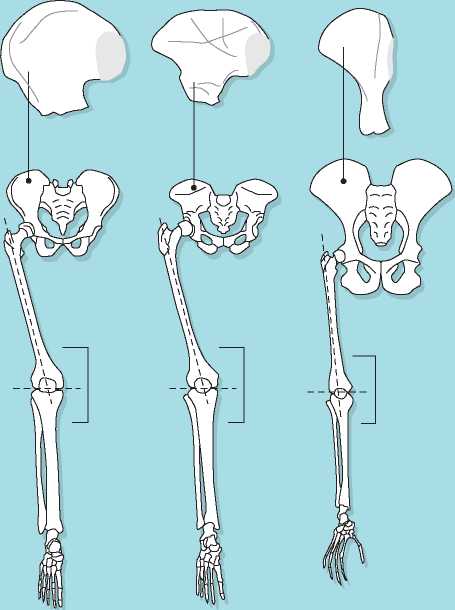
Figure 7.3 Examination of the upper hip bones and lower limbs of (from left) Homo sapiens, Australopithecus, and an ape can be used to determine means of locomotion. The similarities between the human and australopithecine bones are striking and are indicative of bipedal locomotion.
The shape of the pelvis also differs considerably between bipeds and other apes. Rather than an elongated shape following the arch of the spine as seen in chimps, the biped pelvis is wider and foreshortened so that it can provide structural support for the upright body. With a wide bipedal pelvis, the lower limbs would be oriented away from the body’s center of gravity if the thigh bones (femora) did not angle in toward each other from the hip to the knee, a phenomenon described as “kneeing-in.” (Notice how your own knees and feet can touch when standing while your hip joints remain widely spaced.) This angling does not continue past the knee to the shin bones (tibia), which are oriented vertically. The resulting knee joint is not symmetrical, allowing the thigh and shin bones to meet despite their different orientations (Figure 7.3).
Another characteristic of bipeds is their stable arched feet and the absent opposable big toe. The ape big toe is in an abducted position (sticking out away from the midline) while the human big toe is pulled in toward the midline (adducted). In general, humans and their ancestors possess shorter toes than the other apes.
These anatomical features allow paleoanthropolo-gists to “diagnose” bipedal locomotion even in fragmentary remains such as the top of the shin bone or the base of a skull. In addition, bipedal locomotion can also be established through fossilized footprints, preserving not so much the shape of foot bones but the characteristic stride used by humans and their ancestors. In fact, bipedal locomotion is a process of shifting the body’s weight from one foot to the other as the nonsupporting foot swings forward. While the body is supported in a one-legged stance, a biped takes a stride by swinging the other leg forward. The heel of the foot is the first part of the swinging leg to hit the ground. Then as the biped
Continues to move forward, he or she rolls from the heel toward the toe, pushing or “toeing off” into the next swing phase of the stride (Figure 7.4). While one leg is moving from heel strike to toe off of the stance phase,
Abduction Movement away from the midline of the body or from the center of the hand or foot.
Adduction Movement toward the midline of the body or to the center of the hand or foot.
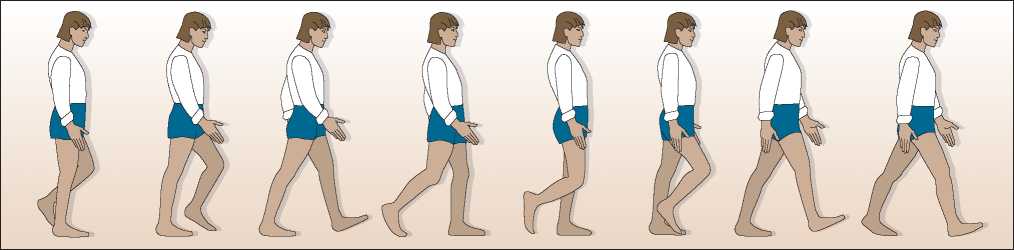
Figure 7.4 The bipedal gait in some regards is really “serial monopedalism" or locomotion one foot at a time through a series of controlled falls. Note how the body's weight shifts from one foot to the other as an individual moves through the swing phase to heel strike and toe off.
The other leg is moving forward through the swing phase of walking.
The most dramatic confirmation of our ancestors’ walking ability comes from Laetoli, Tanzania, where,
3.6 mya, two (perhaps three) individuals walked across newly fallen volcanic ash. Because it was damp, the ash took the impressions of their feet, and these were sealed beneath subsequent ash falls until discovered by chemist Paul Abell in 1978. Abell was part of a team led by British paleoanthropologist Mary Leakey in search of human origins at Laetoli (see Anthropologists of Note). The shape of the footprints and the linear distance between the heel strikes and toe offs are quite human.
Once bipedalism is established in a fossil specimen, paleoanthropologists turn to other features, such as the skull or teeth, so that they can begin to establish relationships among the various fossil groups. Often scholars bring different interpretations to the fossil evidence.
Made famous through the partially complete 4.4-million-year-old skeleton specimen “Ardi,” the genus Ardipithecus has dramatically changed what is known about the earliest bipeds. The genus is actually divided into two species, Ardipithecus ramidus and the older Ardipithecus kadabba dated to between 5.2 and 5.8 mya. The Ardipithecus remains show that some of the earliest bipeds inhabited a forested environment much like that of contemporary chimpanzees, bonobos, and gorillas; these remains were found in fossil-rich deposits along Ethiopia’s Awash

M
The remarkably complete remains of Ardipithecus ramidus have allowed paleoanthropologists to begin to reconstruct the biology and lifeways of the early forest dwelling hominins. When the first “Ardi" remains were discovered in the early 1990s she was placed on a side branch of human evolution but scientific investigation of this remarkable skeleton has lead paleoanthropologists to suggest she is a direct ancestor to the human line.
Anthropologists of Note
Louis S. B. Leakey (1903-1972) ¦ Mary Leakey (1913-1996)

Few figures in the history of paleoanthropology discovered so many key fossils, received so much public acclaim, or stirred up as much controversy as Louis Leakey and his second wife, Mary Leakey.
Born in Kenya of missionary parents, Louis received his early education from an English governess and subsequently was sent to England for a university education.
He returned to Kenya in the 1920s to begin his career there.
It was in 1931 that Louis and his research assistant from England, Mary Nicol (whom he married in 1936), began working in their spare time at Olduvai Gorge in Tanzania, searching patiently and persistently for remains of early human ancestors. It seemed a good place to look, for there were numerous animal fossils as well as crude stone tools lying scattered on the ground and eroding out of the walls of the gorge.
Their patience and persistence were not rewarded until 1959, when Mary found the first fossil. A year later, another skull was found, and Olduvai was on its way to being recognized as one of the most important sources of fossils relevant to human evolution in all of Africa. While Louis reconstructed, described, and interpreted the fossil material, Mary made the definitive study of the Oldowan tools, a very early stone tool industry.
The Leakeys' important discoveries were not limited to those at Olduvai. In the early 1930s, they found the first fossils of Miocene apes in Africa at Rusinga Island in Lake Victoria. Also in the 1930s, Louis found a number of skulls at Kanjera, Kenya, that show a mixture of derived and more ancestral features. In 1948, at Fort Ternan, Kenya, the Leakeys found the remains of a late Miocene ape with features that seemed appropriate for an ancestor of the bipeds. After Louis's death, Paul Abell, a member of an expedition led by Mary Leakey, found the first fossilized footprints of early bipeds at Laetoli, Tanzania.
In addition to their own work, Louis Leakey promoted a good deal of important work on the part of others. He made it possible for Jane Goodall to begin her landmark field studies of chimpanzees; later he was instrumental in setting up similar studies among gorillas (by Dian Fossey) and orangutans (by Birute Galdikas). He set into motion the fellowship program responsible for the training of numerous paleoanthropologists from Africa. The Leakey tradition has been continued by son Richard, his wife Meave, and their daughter Louise.
Louis Leakey had a flamboyant personality and a way of interpreting fossil materials that frequently did not stand up well to careful scrutiny, but this did not stop him from publicly presenting his views as if they were the gospel truth. It was this aspect of the Leakeys' work that generated controversy. Nonetheless, the Leakeys produced a great deal of work that resulted in a much fuller understanding of human origins.
River accompanied by fossils of forest animals. The name Ardipithecus ramidus is fitting for an ultimate human ancestor as Ardi means “floor” and ramid means “root” in the local Afar language.
Now that the spectacular Ardi specimen has been sufficiently analyzed by the team who discovered her, paleoan-thropologists are debating her exact place on the human line. Because the other African apes share a body plan similar to one another, many paleoanthropologists expected the earliest bipeds to resemble something halfway between chimps and humans. Instead, Ardi shows that these forest creatures moved in a combination of ways: They moved
Ardipithecus ramidus One of the earliest bipeds that lived in forested portions of eastern Africa about 4.4 million years ago.
Across the tops of branches with the palms of their hands and feet facing downward, and they walked between the trees on the ground in an upright position. The other African apes, as we saw in previous chapters, knuckle-walk on the forest floor and hang suspended below the branches. In other words, Ardi resembles some of the early Miocene apes more than she does the living African apes.
This calls into question what the last common ancestor of humans and the other African apes looked like. Does Ardi represent the more ancestral form, with the other apes evolving independently after they split from the human line but still converging to the typical African ape body plan? Or does Ardi represent a new body plan, characteristic of the earliest bipeds that evolved away from the African ape plan shared by chimps and gorillas? And what of Ardi’s relationship to the later bipeds? Until fall
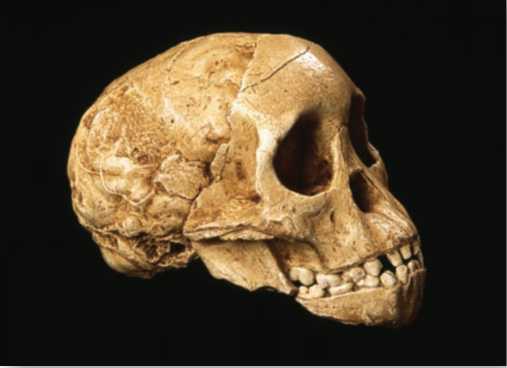
The Taung child, discovered in South Africa in 1924, was the first fossil specimen placed in the genus Australopithecus. Though Raymond Dart correctly diagnosed the Taung child's bipedal mode of locomotion as well as its importance in human evolution, other scientists rejected Dart's claims that this small-brained biped with a humanlike face was a direct ancestor to humans. In the early 20th century, scientists expected the ancestors to humans to possess a large brain and an apelike face and to originate from Europe or Asia rather than Africa.
2009, Ardipithecus was generally considered a side branch on the human evolutionary tree. Now, the international team has proposed that Ardi may be a direct ancestor to the later bipeds, including humans.
The famous Ardi specimen, at 120 centimeters tall and a weight of about 50 kilograms, is comparable in size to a female chimpanzee. The size and shape of this partial skeleton’s brain and the enamel thickness of the specimen’s teeth are similar to chimpanzees as well. Though possessing a grasping big toe, Ardi’s locomotion, unlike that of a chimp, has been reconstructed as bipedal when on the ground.
The Ardipithecus finds, along with the Orrorin and Toumai specimens described in the previous chapter, have begun to provide evidence for the time period before the appearance of the ancient bipeds belonging to the genus Australopithecus. The first representatives of this group were discovered in the early 20th century, long before the majority of scientists were comfortable with the now-accepted notion that humans originated on the African continent.
Most of the early bipeds from the Pliocene are members of the genus Australopithecus, a genus that includes species from East, South, and Central Africa. The name for this group of fossils was coined back in 1924 when the first important fossil from Africa proposed to be a human ancestor came to light. This unusual fossil, consisting of a partial skull and natural brain cast of a young individual, was brought to the attention of anatomist Raymond Dart of the University of Witwatersrand in Johannesburg, South Africa. The “Taung child,” named for the limestone quarry in the South African town of Taung (Tswana for “place of the lion”) in which it was found, was unlike any creature Dart had seen before. Recognizing an intriguing mixture of ape and human characteristics in this unusual fossil, Dart proposed a new taxonomic category for his discovery— Australopithecus africanus or “southern ape of Africa”— suggesting that this specimen represented an extinct form that was ancestral to humans.
Although the anatomy of the base of the skull indicated that the Taung child was probably a biped,
Australopithecus The genus including several species of early bipeds from East, South, and Central Africa living between about 1.1 and 4.3 million years ago, one of whom was directly ancestral to humans.
The scientific community was not ready to accept the notion of a small-brained African ancestor to humans. Dart’s original paper describing the Taung child was published in the February 1925 edition of the prestigious journal Nature. The next month’s issue was filled with venomous critiques rejecting Dart’s proposal that this specimen represented an ancestor to humans. Criticisms of Dart ranged from biased to fussy to sound. Some scholars chastised Dart for incorrectly combining Latin and Greek in the genus and species name he coined. Valid critics questioned the wisdom of making inferences made about the appearance of an adult of the species based only on the fossilized remains of a young individual.
However, ethnocentric bias was the biggest obstacle to Dart’s proposed human ancestor. Paleoanthropologists of the early 20th century expected that the ancestor to humans already had a large brain. Moreover, most European scientists expected to find evidence of this largebrained ancestor in Europe or, barring that, Asia.
In fact, many scientists of the 1920s even believed that the ancestor to humans had already been found in the Pilt-down gravels of Sussex, England, in 1910. The Piltdown specimens consisted of a humanlike skull and an apelike jaw that seemed to fit together, though the crucial joints connecting the two were missing. They were discovered along with the bones of some other animal species known to be extinct. Charles Dawson—the British amateur archaeologist,
The Piltdown forgery was widely accepted as ancestral to humans, in large part because it fit with conventional expectations that the missing link would have a large brain and an apelike face. No one knows with certainty how many of the “Piltdown Gang”—scientists supporting this specimen as the missing link—were actually involved in the forgery. It is likely that Charles Dawson had help from at least one scientist. Sir Arthur Conan Doyle, the author of the Sherlock Holmes detective stories, has also been implicated.

Paleontologist, and practicing lawyer who found these remains—immodestly named them Eoanthropus dawsoni or “Dawson’s dawn man.” Until the 1950s the Piltdown remains were widely accepted as representing the missing link between apes and humans; today they are known as one of the biggest hoaxes in the history of science.
There were several reasons for widespread acceptance of Dawson’s “dawn man.” As Darwin’s theory of evolution by natural selection began to gain acceptance in the early 20th century, intense interest developed in finding traces of prehistoric human ancestors. Accordingly, predictions were made as to what those ancestors looked like. Darwin himself, on the basis of his knowledge of embryology and the comparative anatomy of living apes and humans, suggested in his 1871 book The Descent of Man that early humans had, among other things, a large brain and an apelike face and jaw.
Although the tools made by prehistoric peoples were commonly found in Europe, their bones were not. A few fossilized skeletons had come to light in France and Germany, but they were not at all like the predicted missing link, nor had any human fossils been discovered in England ever before. Given this state of affairs, the Piltdown finds could not have come at a better time. Here at last was the long-awaited missing link, and it was almost exactly as predicted. Even better, so far as English-speaking scientists were concerned, it was found on English soil.
In the context of the evidence available in the early 1900s, it was easy to accept the idea of an ancient human with a large brain and an apelike face. Fortunately, the self-correcting nature of science has prevailed, exposing the Piltdown specimens as a forgery. The discoveries— primarily in South Africa, China, and Java—of fossils of smaller-brained bipeds from the distant past caused scientists to question Piltdown’s authenticity. Ultimately, the application of the newly developed fluorine dating method (described in Chapter 5) by British physical anthropologist Kenneth Oakley and colleagues in 1953 proved conclusively that Piltdown was a forgery. The skull, which was indeed human, was approximately 600 years old, whereas the jaw, which proved to be from an orangutan, was even more recent. Finally, Dart and the Taung child were fully vindicated.
Today, genetic and fossil evidence indicates that the human evolutionary line begins with a small-brained bipedal ape from Africa. Numerous international expeditions—including researchers from Kenya, Ethiopia, Japan, Belgium, Great Britain, Canada, France, Israel, the Netherlands, South Africa, and the United States—have scoured East, South, and Central Africa, recovering unprecedented amounts of fossil material. This wealth of evidence has allowed scientists to continually refine our understanding of early human evolution. Even though debate continues over the details, today there is widespread agreement over its broad outline. Each new discovery, such as the Ardi skeleton, confirms our African origins and the importance of bipedalism to distinguishing humans and their ancestors from the other African apes.
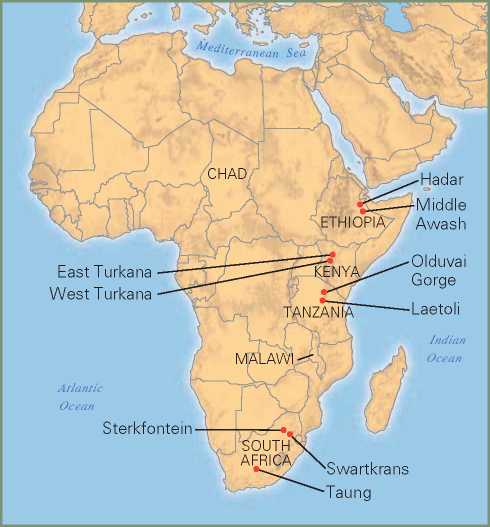

Figure 7.5 Australopithecine fossils have been found in South Africa, Malawi, Tanzania, Kenya, Ethiopia, and Chad.
In the Miocene the Eurasian and African continents made contact at the eastern and western ends of what now is the Mediterranean Sea. As these landmasses met, “rifting" also occurred, gradually raising the elevation of the eastern third of Africa. The dryer climates that resulted may have played a role in human evolution in the distant past. This rifting also gives us excellent geologic conditions for finding fossils today.
 World History
World History









