



1 Provenience. Artifacts from Component A were recovered at a depth of about 9-10 m from the present ground surface. They were found in good stratigraphic and spatial association with the faunal remains. Out of a much larger number of pieces recovered by Lbova, we examined 552 (Table A1.1, site 8). These are about half the total studied by Germonpre and Lbova (1996), whose taphonomic work we will be comparing wherever possible. We could see no reason not to pool all of the Component A pieces, and excluded Components B and C because of apparent stone tool differences and small sample sizes. While we recorded all of the provenience information on each individual piece of bone - such as year, component, square - in some cases year or section were not identified. Informal inspection of the provenience information failed to turn up any obvious spatial or temporal pattern for any of our variables.
2 Species. We examined 552 Component A pieces, with gazelle being the most frequently recognized species (57.1%) (Table A1.2, site 8). There were small amounts of horse (4.7%), and even lesser amounts of bison, camel, rhinoceros, and wolf. Less-specific pieces and those that were unidentifiable made up the remainder. Significantly, there was no direct or indirect evidence of hyenas. The range of fauna analyzed by Germonpre and Lbova (1996) is mentioned above.
A statistical comparison of the frequencies of our identified species with the 2009 pieces (all archaeological components) studied by Germonpre and Lbova (1996:40) produced a very significant difference (x2 = 70.0, 12 d. f. , p<0 .001). The difference was caused by:
(1) our under-identification of horse, because we did not study most unmodified whole bones; (2) we identified many more examples of gazelle (Procapra), attributable to Ovodov’s many years of paleontological experience; and (3) we had many fewer unidentified pieces, again probably due mainly to Ovodov’s experience plus our minimal size limitation that excluded fTom study pieces smaller than 2.5 cm in diameter. Given our different study objectives (perimortem taphonomy versus standard taphonomy) and sampling methods, the significant difference is due mainly to our recognition of more gazelle remains.
There are two obvious differences in the species found at Kamenka and our other sites. Most notable is: (1) the absence of gazelle in all other sites except Varvarina Gora, which is only 17 km from Kamenka and was occupied during a similar time period; (2) hyenas were absent at Kamenka.
3 Skeletal elements. Our element sample size was 552 pieces (Table A1.3, site 8). Pieces identifiable as long bone but not specific bones were the most numerous (12.7%), followed by pieces of rib (11.4%). Both classes are readily identifiable, but then so are cranial elements, which taken together make up 12.5% of our total. There were no conspicuously missing elements, although given the ease with which vertebrae can be recognized, the large number per individual, and their generally good archaeological preservation, there seems to be a deficiency in their expected occurrence. Fragmented skeletal elements are generally easier to identify than are species.
The skeletal element comparison with Germonpre and Lbova (1996:41) produced a highly significant difference (x2 = 478.1, 18 d. f., p <0 .001). The main difference is due to Germonpre and Lbova’s identification of many more long bones (“unidentified diaphyses”) and unidentifiable pieces. The difference between our respective unidentifiable pieces is attributable to Ovodov’s years of experience, and our exclusion of pieces <2.5 cm. However, as with our respective species counts, overall the element identifications are reasonably similar.
On the whole, Kamenka displays nothing out of the ordinary with respect to its skeletal element frequencies compared with our other sites. Only one oddity stands out, which was the occurrence of four pieces of penis bone (0.7%). This element occurred elsewhere only in nearby Varvarina Gora, and in the faraway mountainous Altai caves of Ust-Kan and Razboinich’ya Cave. The latter has the highest frequency of penis bones in our entire series - 5.4%. According to Davis (1987:44), male equids, rodents, and carnivores have penis bones.
4 Age. Our sample size for age assessment was 552 pieces (Table A1.4, site 8). Adults were represented about five times more often than sub-adults, assuming that the pieces of unknown age were in roughly the same 5:1 ratio. One Procapra ulna with osteophytic proliferation around the proximal end and a healed antemortem fracture of the shaft obviously belonged to an adult. Germonpre and Lbova do not report on age.
Comparing our other human-caused bone samples of comparable size with Kamenka, Mal’ta, Varvarina Gora, and Volchiya Griva have similar adult/sub-adult ratios. Most of the others have far fewer sub-adults - for example, Bolshoi Yakor, Denisova, Kaminnaya, Ust-Kan, Ust-Kova, etc. This age ratio difference is not solely attributable to age-dependent bone hardness and calcification, because preservation is good in these cave and open sites, among others. Seasonal hunting techniques, early-in-life deaths due to health and nutrition (Volchiya Griva), and heavier bone processing of younger animals by humans and carnivores alike are potential factors influencing the inter-site age ratios.
5 Completeness. We studied 552 pieces for completeness (Table A1.5, site 8). Our whole pieces (8.7%) are mainly those that had some kind of processing stigmata. Those with one remaining anatomical end make up 37.5%. Pieces lacking both anatomical ends were in the majority (53.8%).
By combining our two incomplete categories we could statistically compare “breakage” occurrence with Germonpre and Lbova. The comparison showed no significant difference (X2 = 0.2, 1 d. f, p>0.5). Hence our respective whole/breakage ratios are effectively the same, namely 9.0% whole, 91% broken. There is nothing especially unusual about Kamenka compared with our other assemblages. A frequency of roughly 50% lack of both anatomical ends lies near the mean of our other sites.
6 Maximum size. The average maximum diameter for the 550 pieces that were either whole or if broken were unaffected by postmortem breakage is 6.5 cm, and the range is 2.9 cm to 20.7 cm (Table A1.6, site 8). While Germonpre andLbova (1996:42) provide a percentage histogram for fragment length, we are unable to convert it to precise values for statistical comparison. The shape of the histogram is highly skewed, but the median is clearly 3-5 cm, with what we estimate to be about 55% of their unstated total. The histogram shows that their second most frequent class had a class interval of 0-2 cm, and a frequency of about 20%. Hence, their median and our mean values are probably not significantly different, given our different criteria for measurement inclusion.
The Kamenka mean piece size is, as was the case for completeness, similar to the combined mean (9.0 cm) for our other assemblages. Comparing the Kamenka size range values with undamaged long-bone lengths provided by Vera Gromova (1950: table 27), shows that the former upper limit is about the same as the values for Saiga antelope, but smaller than those of sheep (Gromova has no information on gazelles).
7 Damage shape (Figs. 3.28-3.30). Damage form was assessed in all 552 Kamenka pieces (Table A1.7, site 8). Most frequent are long bone fragments (36.9%), long bone splinters (11.4%), and long bone flakes (11.2%). Overall, damage-form occurrence correlates, of course, with skeletal element occurrence. Long bone mass (volume or weight) probably exceeds that of other categories such as crania, vertebrae, flat bones, and irregularly shaped bones.
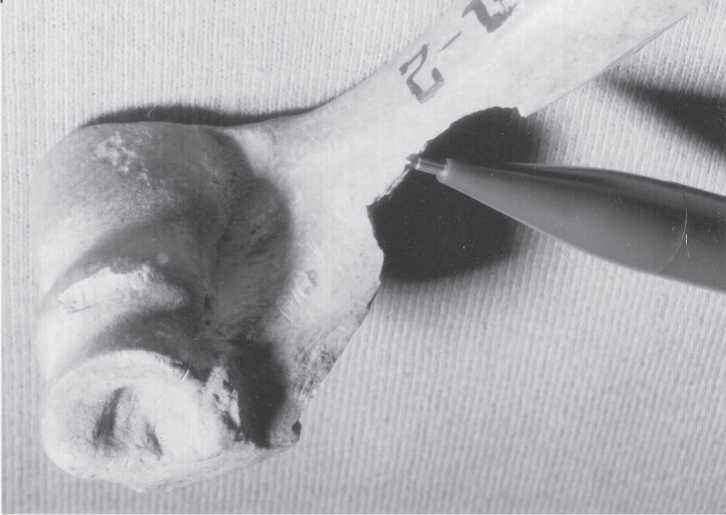
Fig. 3.28 Kamenka bone damage. A Procapra humerus with a big chop notch (at pencil) and cut marks on joint. Fragment length, 7.0 cm. Layer A (CGT neg. MBSC 7-4-03:5).
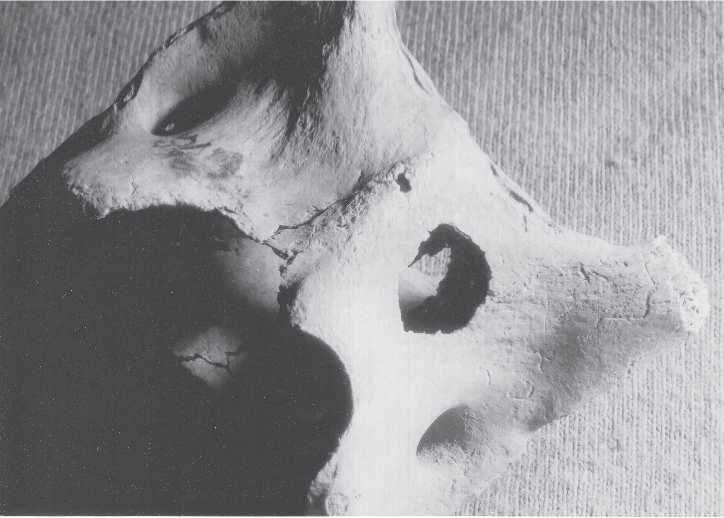
Fig. 3.29 Kamenka bone damage. A punched hole 1.0 x 0.8 cm in a Procapra first cervical vertebra, 5.0 cm maximum diameter. Layer A (CGT neg. MBSC 7-4-03:2).

Fig. 3.30 Kamenka bone damage. Procapra humerus with cut marks near joint. Side B. Actual width of image is 3.3 cm. Layer A (CGT neg. MBSC 7-4-03:5).
Germonpre and Lbova (1996:42) consider damage form as a variable they call “traces.” Traces include gnawing marks, root marks, burning, cut marks, percussion marks, cancellous blocks, and bone splinters. They recognized that Kamenka had a large number of narrow and variably long pieces that look like splinters. They report that 10.5% of their 1928 pieces are splinters (202 splinters). As mentioned, we recognized 11.4% of our total (552) as being splinters. There is no significant difference in our findings (x2 = 0.35, 1 d. f., p>0.05).
These authors identified a Kamenka damage form they called “cancellous blocks,” which tallied at 3.3% (64/1928). We saw several of these but did not count their occurrence. However, cancellous blocks most likely are derived in the main from vertebral bodies, ends of long bones, and the cancellous bone around the pelvic acetabulum (“pelvis center”). Combining our occurrences of vertebral bodies (1.6%), long bone epiphyses (0.4%), and pelvis centers (2.7%) totals 4.7%, a value similar to Germonpre and Lbova’s 3.3%. Among rare damage variants, we came across a single case of both horn cores of an adult Procapra cranial vault having been pounded off at the horn-vault junction, the purposes of which we speculate had been to get the 9.8 cm wide vault piece into a cooking bag, or to use the horns for some sort of tool, or both.
Another rare damage variant was the occurrence of a seemingly punched 1.0 x 0.8 cm circular hole in the corpus of an adult Procapra’s first cervical vertebra.
In comparison with our other assemblages, Kamenka does not stand out as unusual in any of the damage types. While the Kamenka percentage of long bone splinters is among the assemblages with the most splinters, sites such as Bolshoi Yakor (25.1%), Dvuglaska (15.1%), and Ust-Kan (16.9%) have more.
8 Color. Color was assessed in all 552 pieces (Table A1.8, site 8). Most of the pieces (98.9%) were scored as ivory in color. Postmortem breakage shows that Kamenka bone coloring is almost entirely surficial. The remaining 1.1% were classified as brown in color. These were pieces that probably had been burned, because the color seems to have been present in the interior of each piece, judging fTom a few tiny breaks we made on existing fractured edges. We use color as the main identifier of burning. In addition, pieces that we classified as burned had almost no polishing, which we suggest was due to: (1) the pieces having fallen accidentally into a fire before they could acquire polishing; or (2) burned pieces not retaining polishing through time as well as unburned pieces. Hence, a lack of polishing reinforces our view that brown and black colored pieces are the result of burning.
We ignore surficial staining in our color identification, whereas Germonpre and Lbova (1996:41) considered staining, reporting that “colour varies from a dark greyish brown to a lighter brown, but some bones exhibit a light grey hue.” One of their “traces” categories is burned fragments, which were rare (1.3%) and small (<5.0 cm long). We agree that most of the burned pieces are smaller than 5.0 cm, although we did identify three probably burned pieces that varied from 7.0 cm to 10.9 cm in length. A comparison of our respective burning frequencies (ours: 1.1%) shows no significant difference (x2 = 0.23, 1 d. f.,p >0.05).
Compared with our other assemblages, Kamenka is generally without distinction, unless only open archaeological sites are compared. Then, Kamenka does stand out because of its very low amount of deep discoloration. Faunal remains from the open sites at Bolshoi Yakor and Ust-Kova have much lower frequencies of ivory colored pieces. We do not understand the cause of this difference. Lbova (personal communication, July 2003) suggested that rapid burial by shifting sand may have played an important role in these open-site color differences.
9 Preservation. All 552 pieces were assessed for quality of preservation (Table A1.9, site 8). Most pieces were ivory hard and excellently preserved (94.4%) - they could not be scratched with a fingernail. The remainder were chalky or intermediate, that is both chalky and ivory in hardness. One rare quality variant, a Procapra rib fragment, had a high-quality ivory external surface and a chalky and root-track-riddled lung-facing surface. Presumably the burial position of this piece had the chalky side facing upward, toward the ground surface.
Germonpre and Lbova (1996:41) similarly found that preservation was generally excellent. Using a different method to assess preservation, they found 94.1% had no or only minor weathering. While we are unsure of all the ways bone is degraded to a chalky state - surface weathering being certainly an important factor - we are in complete agreement with their assessment of preservation, including their suggestion that the Kamenka Component A assemblage was on the whole exposed on the ground surface or near the surface for only a short time before deeper burial. We would add that shallow frozen ground would have aided preservation before and after the relatively short summers.
Inspection of bone quality in each of our other assemblages (Table A1.9, site 8) quickly reveals that most cave deposits such as Denisova and Kaminnaya have excellent bone preservation, whereas open sites have assemblages with greater amounts of chalky bone (Kara Bom, Malaya Seeya, Mal’ta, and so forth). But there are exceptions other than Kamenka. These include Krasny Yar and Kurla (while Boisman is an open site with very little chalky bone, it is much younger than all the rest, and it contains large amounts of shell that helps buffer bone-damaging soil acids). It is also evident that open sites containing mainly mammoth remains (Shestakovo and Volchiya Griva) exhibit very poor preservation. Factors involved in this poor preservation probably include bone characteristics such as the relatively thin cortex of mammoth skeletal elements, the longer periods of time these very large bones remain unburied due simply to their size, and the contextual conditions of their locations that might include only modest sources of windblown or water-deposited overburden, trampling, and other physical and mechanical hazards to preservation.
10 Perimortem breakage. All 552 pieces could be assessed for perimortem breakage (Table A1.10, site 8). Most pieces of the Kamenka assemblage had perimortem breakage (94.7%). When this value is coupled with piece size (Table A1.6, site 8) it is clear that very extensive carcass processing took place in the Kamenka camps. This severe bone reduction and fragmentation has been viewed by a number of workers as indirect evidence of cooking for the purpose of extracting as much nutrient as possible from a carcass, especially fat-rich, high-energy marrow found in the interior of many skeletal elements. As we will mention time and again, we propose that cooking was done in hide or other sorts of containers holding water, fTesh bone fragments with adhering flesh, globs of marrow, and heated stones, i. e., bag-boiling. An animal’s gutted thorax and abdominal cavity could also serve as a container for the stone-boiling of itself.
A comparison of our perimortem breakage findings with those reported by Germonpre and Lbova (1996:41) shows no significant difference (x2 = 0.21, 1 d. f.,p>0.05).
The Kamenka perimortem breakage incidence is much like that of our other assemblages, except for the lower occurrence in the open paleontological sites (Krasny Yar, Shestakova, Volchiya Griva). In contrast, the paleontological cave sites such as Razboinich’ya have a great deal of perimortem breakage.
11 Postmortem breakage. All 552 Kamenka pieces were examined for postmortem breakage (Table A1.11, site 8). Identifiable postmortem breakage is largely associated with general excavation problems and specifically the difficulty of extracting crumbly mammoth bone fTom its matrix. Postmortem breakage is rare (1.1%) in the Kamenka assemblage. This is attributable to high-quality excavation, excellent preservation, a finegrained homogeneous matrix, and the absence of easily fragmented mammoth remains in Component A. However, modified mammoth tooth that had been made into a bracelet or diadem artifact was recovered (Lbova 2000:176), and one non-artifact piece of mammoth was identified by Germonpre and Lbova fTom Component B. These authors report finding no examples of “recent” bone fractures, which we presume to mean postmortem breakage.
Compared with our other assemblages, Kamenka has a very low frequency of postmortem breakage. Of the fully archaeological sites, only Kurla has a lower value.
12 End-hollowing. Only 240 Kamenka pieces could be evaluated for end-hollowing due to the fact that an anatomical end is generally required to assess the occurrence of this variable. Of these pieces, only 0.8% exhibit end-hollowing. These two pieces also have polishing on both ends, and one has tooth dints and small notches in a perimortem fracture. We consider end-hollowing to be a useful but not infallible criterion for carnivore damage.
Germonpre and Lbova report that carnivore gnawing is rare in their study of the Kamenka assemblage (n = 26,1.6%); however, they do not report how they recognized it. Since one of the primary goals of our study is to define the signature of carnivore perimortem damage, we suggest that our respective studies are very similar in this inference, but ours is based on the use of multiple variables, end-hollowing being just one. Hence, we have 31/550 pieces (5.6%) that exhibit multiple criteria (excluding perimortem breakage), which we believe represent damage done by carnivores. This multiple-variable issue will be discussed in detail later.
End-hollowing is much more frequent in several of our other assemblages, especially those with direct or strong indirect indications of hyena presence. A return to Table A1.2 will show which sites had hyena bones (direct evidence), while Table A1.21 provides the frequencies for bone fragments with acid erosion (indirect evidence). It is well known that even domesticated carnivores sometimes swallow pieces of bone. If not regurgitated these pieces of bone can show some degree of digestive erosion (Davis 1987:26-27). The occurrence of end-hollowing and a few pieces of acid-eroded bone, plus other criteria of carnivore bone damage, strongly indicates that carnivores were utilizing some of the remains of butchering and cooking left by the human inhabitants of Kamenka. Ovodov’s identification of one wolf bone in the Kamenka assemblage points to that species as
Having been responsible for the end-hollowing. Germonpre and Lbova also identified a bone of a large carnivore as that of a lion. Both species may have occasionally scavenged the refuse of the seasonally departed human hunters and their families, although modern wolves and hyenas are better known for scavenging decayed and dried carcasses than are lions (van Lawick and van Lawick-Goodall 1970).
13 Notching. We could assess notching on 549 pieces, ofwhich 9.8% were so modified (Table A1.13, site 8). Notching can be done by hyenas, as testified to by their common occurrence on bones recovered from Razboinich’ya Cave, and by humans, where some abrasions caused by a hammer stone or anvil stone are in direct association with a notch. The majority of the notched Kamenka pieces had only one notch (6.0%). There were no pieces with more than four notches. Of the notched pieces there were 23 (42.6%) that had neither carnivore nor chop marks. There were 17 (31.5%) notched pieces with some carnivore damage (end-hollowing, tooth scratches, tooth dints, heavy polishing, erosion) but no chopping. Lastly, 14 (25.9%) notched pieces had only chop marks. There were no notched pieces with both chop marks and carnivore damage. Hence, notching by itself is not a sure sign of either hyena or human bone processing at Kamenka.
Germonpre and Lbova (1996:43) found a small number of percussion marks (n = 21; 1.1%) on long bone shafts, most of which were associated with spiral fracturing. They attribute these marks to human bone processing for marrow extraction when found in association with marks attributed to knives, burins, and scrapers. Some of their percussion marks could be due to carnivore notching, just as some of our notching appears to have been done by humans. In our early examinations of the Razboinich’ya assemblage we soon abandoned spiral fracturing as a uniquely human-caused attribute because there were so many pieces that had to have been spirally fractured by hyenas.
The occurrence and multiplicity of notching at Kamenka is not exceptional compared with our other assemblages (Table A1.13). Notching is most frequent in those sites with direct or strong indirect evidence of hyenas or other carnivores having used the caves, including Razboinich’ya, Proskuryakova, Maly Yaloman, Okladnikov, and others. Notching is least fTequent in open archaeological sites.
14 Tooth scratches. We could assess tooth scratching in 541 pieces (Table A1.14, site 8). Only 2.8% exhibited scratching. There is a wide range in the number of scratches per piece, with eight pieces having more than seven scratches. The largest number of scratches on a single piece is 30. While one or even possibly two scratches might have been caused by some agency other than carnivore chewing - such as trampling - several scratches on a piece are most probably the result of carnivore chewing. This is especially so when their occurrence on a piece is substantially parallel, variable in width and depth, and ends in a kind of squiggle. Looking at co-occurring damage in the eight pieces with more than seven scratches we find that all but one have tooth dints as well. The one piece with 40 scratch marks has 25 tooth dints. At least one of the eight had very fine scratches, suggesting a scavenger of fox-like size.
Comparing only our tooth scratching category with the “gnawing marks” category of Germonpre and Lbova (1996:42), we found no significant difference between our respective frequencies (x2 = 0.23, 1 d. f., p>0.05).
Compared with our other assemblages, Kamenka tooth scratching is at the low end of the range (Table A1.14), which generally characterizes open sites. Cave sites have the highest frequencies of tooth scratching.
15 Tooth dints. We could assess 544 Kamenka pieces for tooth dints (Table A1.15, site 8). Like tooth scratching, tooth dinting is also uncommon (3.3%). The range of dints per piece is also wide. Both are strongly related, but not so much as to have to look upon them as a single variable. While there are 14 Kamenka pieces with both dinting and scratches, there are four pieces with only dinting, and two pieces with only scratches. As with tooth scratches, Kamenka, being an open site, has a low frequency of pieces with dints, whereas cave sites generally have higher dint frequencies.
16 Pseudo-cuts. We could assess 546 pieces for pseudo-cuts (Table A1.16, site 8). Pseudo-cuts were very rare (0.4%). When present there were only one or two pseudo-cuts on each of two pieces. Both pieces also had tooth scratches and dints. One of the pieces also had 20 tooth scratches and five tooth dints, the very fine and delicate size of both classes suggestive of chewing by a fox-sized carnivore. Pseudo-cuts exist as a variable because they were first identified in the faunal remains from Razboinich’ya Cave. The cave had no evidence of having ever been inhabited by prehistoric humans. Hence, what appeared to be cut marks had to have been caused by carnivore bone chewing. While an occasional piece with actual cut marks might have been picked up by hyenas scavenging one or another of the human sites in the valley below, and brought back to Razboinich’ya, we doubt this ever happened because pieces with burning and cut and chop marks at joints were not found in the Razboinich’ya assemblage. Abrasions were, however, present, but they could have been due to trampling or skittering of bone on the gritty limestone cave floor.
The rare pseudo-cut marks at Kamenka are similarly rare in other open sites. They are generally more common in the cave sites.
17 Abrasions. We were able to assess 544 pieces for abrasions (Table A1.17, site 8). Only one piece (0.2%) had abrasions. This was a piece of a horse long bone with a set of ten abrasion grooves. The piece also had nine cut and two chop marks.
Abrasions are generally rare in all of our assemblages. It would seem that most bone breakage was done with the aid of non-abrasive hammers and anvils, such as another piece of bone, antler, or wood.
18 Polishing. We were able to assess 540 pieces for polishing (Table A1.18, site 8). Of these, 90.0% had polishing on one or both ends (24.8%), in the middle (4.0%), or on the ends and middle (61.3%). We are uncertain how this polishing forms. It is possible it develops by some natural chemical or physical post-depositional process. On the other hand, prehistoric polishing of mainly ends of human bone fragments has been proposed by White (1992), and verified by Turner and Turner (1999), to have likely occurred when freshly smashed pieces of bone were stirred in a water-filled pottery cooking pot with a slightly gritty interior. This treatment helps extract marrow fat as well as loosen and cook adhering pieces of muscle tissue. Perhaps the Kamenka polishing happened when pieces were abraded by heated stones placed in water-filled hide or tightly woven fiber bags for the same marrow-extracting objective.
Compared with our other assemblages, the frequency of Kamenka polishing is at the higher end of the range, but not exceptionally so (Table A1.18). Because polishing is found in our hyena cave assemblages, carnivore bone chewing must be another mechanism producing this condition. This is especially so when polishing of a piece is extensive and severely rounded.
19 Embedded fragments. We were able to assess 547 Kamenka pieces for embedded fragments (Table A1.19, site 8). Of these, 1.5% exhibited this condition. In our preliminary work we found embedded fragments largely associated with carnivore damage, and most notably in large tooth dints. In the course of time we found a few instances of embedded fragments associated with what were unquestionably hammer percussion indentations and notches. Kamenka has three pieces with embedding but no chop or cut marks, four pieces with embedding and chop or cut marks, and one piece with embedding and digestive erosion. The latter, while definitely eroded, lacks the greasy or slippery feel of hyena stomach bones, which allows for the possibility for some form of soil-dependent damage.
Compared with our other assemblages the frequency of Kamenka pieces with embedded fragments is relatively low (Table A1.19). The cave sites, as expected, have relatively more embedding, but there are notable open site exceptions such as Kurla and Ust-Kova.
20 Tooth wear. We had only 12 tooth-bearing pieces in the Kamenka series, hardly enough for any comments other than very limited suggestions about age. The greater the amount of wear in an assemblage indicates a generally older series of individuals than in an assemblage with less wear. Kamenka seems to have few young individuals at 8.3% (Table A1.20, site 8).
Had the very small sample sizes in Table A1.20 been larger, we would feel much more secure in pointing out that sites such as Boisman II and Kaminnaya, and others, regardless of antiquity differences, apparently have more young individuals than did Kamenka.
21 Acid erosion. There were 550 pieces that could be assessed for the occurrence of digestive damage (Table A1.21, site 8). Kamenka has a low frequency of erosion (1.3%), and of these seven pieces, six have only limited erosion and lack the slippery-feeling and scoured surface of stomach bones associated with hyena deposits. It may be that the seven stomach bones represent less-digested pieces swallowed by wolves, other carnivores, or perhaps even the Kamenka humans themselves. The sizes of these pieces were: 2.5, 3.0,4.3,4.5, 4.5, 5.5, and 5.7 cm, the diameters of which are all well within the range of stomach bones found in the hyena caves. If these weakly eroded pieces represent regurgitation that limited their time in the digestive track, we would expect the larger ones, not all seven, to have had such short acid exposure. Studies of dried human feces from various archaeological localities have shown considerable ingesting of bone fragments by humans (see, for example, Clary 1984, Heizer 1967).
Kamenka is on the low end of the range for the occurrence of digestive damage (Table A1.21). The highest frequencies are associated with hyenas.
22-24 Rodent gnawing, insect damage, and human bone. No examples of these variables were found in the Kamenka assemblage we examined (Tables A1.22, site 8 to A1.24, site 8).
25 Cut marks. A total of 546 pieces could be evaluated for cut marks. Of these, 9.5% had one ormore cut marks (Table A1.25, site 8). Most frequent were pieces with only one incision (3.8%). Of the pieces with the greatest number of cut marks, one is a long bone fragment 11.4 cm in length with 16 cut marks; a sphnter 8.5 cm long has 20 cut marks that may have been produced during excavation; a 7.5 cm long distal portion of a Procapra humerus has 24 cut marks; and a 12.8 cm horse long bone with 64 cut marks is of special interest because the cutting tool left multiple parallel lines as if a unifacially chipped flake had been drawn across the surface like a scraper or plane rather than like the slicing action of a knife.
A comparison of our cut mark frequency with that of Germonpre and Lbova shows a very significant difference (x2 = 111. 3, 1 d. f., p <0 .001). We identified about ten times more pieces with cut marks (52/ 546) that did Germonpre and Lbova (19/1928). The difference is attributable to the many more years of experience in perimortem taphonomy by Turner (Turner and Turner 1999; and elsewhere) than by Germonpre and Lbova.
Compared with our other assemblages, the occurrence of Kamenka cut marks is near the middle of the range. The Kamenka frequency (9.5%) is much like that of nearby Varvarina Gora (8.8%).
26 Chop marks. The Kamenka sample had 548 pieces that could be assessed for chop marks. Ofthese, 8.6% had chop marks (Table A1.26, site 8). The majority (6.2%) ofthese had only one chop mark. Compared with our other assemblages Kamenka is near the middle of the range for chop mark frequency.
 World History
World History









